ヒトの遺伝情報-染色体 (クロモゾーム)
生物は親から子へ膨大な量の遺伝情報を伝えてゆきます。この遺伝情報は細胞の核内にある染色体(Chromosome)の中にまとめられています。
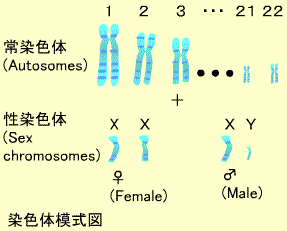
ヒト細胞の核内には23対の染色体があり、これらのうち22対は母親と父親からそれぞれ1つずつ子供に伝えられたものが対になっていて常染色体と呼ばれます。常染色体は長いものから順に1番から22番まで番号がついています(21だけは22番より短くなっています)。残り1対の染色体は性染色体と呼ばれ、子供の性別を決定します。

性染色体異常:
染色体の異常で性染色体の数が変わることがあります。クラインフェルター症候群では2つのXと1つのYをもつ男性が生まれます(XXY)。またターナー症候群では1つのXだけでYを持たない女性が生まれます(XO)。
常染色体異常:ダウン症候群(Down syndrome):
21番染色体が3つあり(トリソミー)、心疾患や神経症状を伴います。
性決定
X染色体の数は性別決定に関係なく、Y染色体を持つかどうかが性別決定に重要なことがわかっています。この性別決定にはY染色体の末端付近にあるSRYという遺伝子が関与しています。
SRY遺伝子の塩基配列は性別判定の検査にも利用されています。
(例外として染色体のクロスオーバーによってSRYがXに移動するとXXの子孫でも男性的な特徴をもち、また何らかの原因でSRYをYから失うと子孫はXYでも女性的な特徴を持ちます。)
X染色体の不活性化 (X inactivation)
染色体上の遺伝子は発現して蛋白となりますが、X染色体を2つ持つ女性がXを1つしか持たない男性に比べて2倍量のX染色体遺伝子を発現することはありません。これは受精後の初期段階で女性の2つあるX染色体の片方が不活性化され、この染色体からの遺伝子発現が抑制されるからです。これをX inactivationと呼びます。
X染色体が2個以上あっても1つをのぞき、他はすべて不活性化されます(実際にはX inactivationを逃れる遺伝子もあります)。不活性化されたX染色体は顕微鏡でみるとバール体(Barr body)と呼ばれるかたまりに見えます。
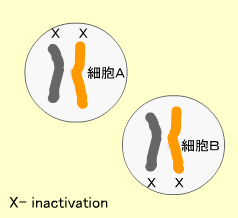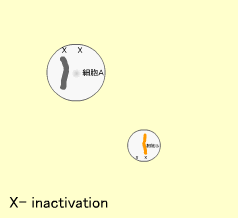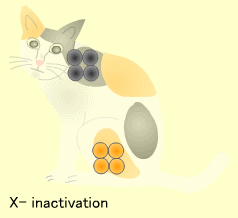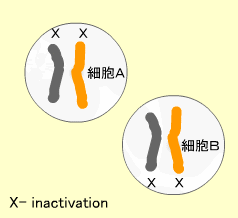
どちらのX染色体が不活性化されるかはランダムですが、一度不活性化されるとその後の細胞分裂でもこの不活性化は保たれます。よって体のある部分では父親由来、他の部分では母親由来のXが不活性化された細胞集団が存在するようになります。
X染色体の不活性化にはXistという遺伝子がかかわっています。この遺伝子の発現によって片方のXが不活性化されます。
インプリンティング (Genomic Imprinting)
常染色体上の遺伝子は対になっているそれぞれ(母親と父親由来)に同じ遺伝子コピーを持っており(biallelic)、それぞれが発現して蛋白となっていますが、例外的に父親または母親由来の片方しか発現しない遺伝子があることが知られています。
これらの遺伝子は遺伝的に刷り込み(インプリンティング)されており、父親か母親由来のどちらだけが発現して蛋白になるかが決まっています。インプリンティングにはDNAのメチル化が関係しているといわれています。
- IGF-IIという遺伝子は父親からのもののみ発現し、母親からのものは発現しません(脳内では両方からのコピーが発現しています)。
- プレイダー・ウイリィー症候群(Prader-Willi syndrome)とエンジェルマン症候群(Angelman syndrome):
この2つの病気は全く異なった症状を示しますが、ともに第15染色体の部分欠損(15q11-13)が関係しています。父親由来の染色体からこの部分が欠損するとプレイダーウイリィー症候群、逆に母親由来の染色体部分が欠損するとエンジェルマン症候群になります。 - インプリンティングと体臭:
女性がある男性の体臭を不快に感じるかどうかは父親由来のHLAのタイプによります(Nature Genetics, 2002)。
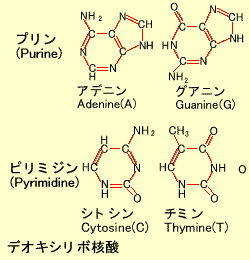
DNA (Deoxyribonucleic Acid)
染色体は巨大な糸巻きのようなもので、ヒストンというタンパク質のまわりにDNA鎖が巻き付いています。DNAはアデニン(A)、グアニン(G)、シトシン(C)、チミン(T)の4つの塩基(デオキシリボ核酸)からなります。これらの塩基はその構造からアデニンとグアニンはプリン(Purine)、シトシンとチミンはピリミジン(Pyrimidine)と呼ばれます。
タンパク質をコードする遺伝子はこれらの4つの塩基が正しい順番でならんだ構造をとっています。このDNA鎖はひとでは全体で30億塩基対ありますが、そのほとんどの配列はすでに決定されています。
DNAのメチル化 (Methylation)
ほ乳類のシトシン (C) の3%はメチル化(CH3が付く)されています。このメチル化はCpGというCとGが連続した場所に多くみられ,CpGの80%ぼどがメチル化されています。ゲノム上でCpGが高密度に存在する場所をCpGアイランド(Island)といい、遺伝子発現の調節、癌、インプリンティング等に関係していると言われています。
一般的にCpGアイランドのシトシンはあまりメチル化されておらず、それ以外の部分のCpGはメチル化されています。おもしろいことに酵母、線虫、ドロソフィラ(ハエ)のゲノムは全くメチル化されていません。通常の塩基配列決定法ではメチル化は検出できないので、メチル化に敏感な制限酵素や化学的な方法で検出する必要があります。
DNAのメチル化部位は遺伝子変異の好発部位になります。これはメチル化されたシトシンはチミン(T)に化学的変化を起こしやすいためです。ひと個人の染色体上には1000塩基対に1つ遺伝子配列の違いがあると言われています。これを1塩基多型性(Single nucleotide polymorphism: SNP)と呼びます。SNPはひと個人間の薬の効き方の違い等に関係していると言われています。また、乳ガンに関係した遺伝子BRCA1にはこのメチル化部位が変異を起こしている例が報告されています。
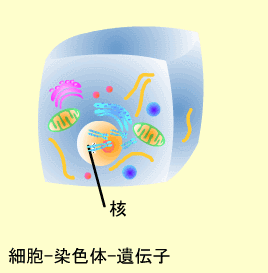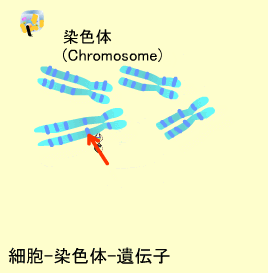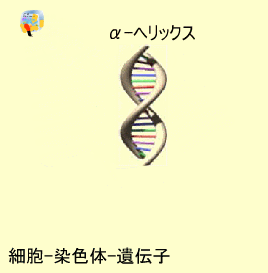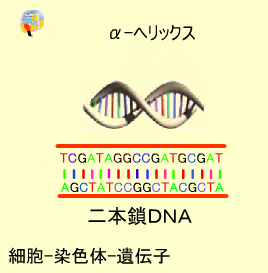
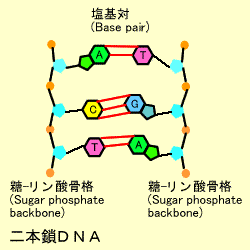
二本鎖DNA
DNAの鎖は2本がらせん階段のようにペアになった構造をしています。これをダブルへリックス(Double Helix)といいます。この際、アデニン(A)とチミン(T)、シトシン(C)とグアニン(G)はお互いに対になって結合しています。これを塩基の相補性(complimentary)といいます。
言い換えるとプリンはいつもピリミジンと向かい合っています。このへリックス構造は塩基10個で1回転します。アデニンとチミンの間は2つ、シトシンとグアニン間には3つの水素結合があり、二本鎖構造を保っています。
DNAフィンガープリンティング (DNA Fingerprinting)
染色体DNAの特定の部分を比べると個人によって違いのある配列があります。また染色体上に多数存在する反復配列(リピート)、たとえばVNTR(Variable Number of Tandem Repeat:15-70塩基程度の長さ)やSTR(Short Tandem Repeat:2-4塩基)なども個人間でそのリピートの数に大きな変異があります。
これらの遺伝情報の違いは最初、DNAを制限酵素で切断するRFLP:Restriction Fragment Length Polymorphism法によって検出されていましたが、最近は、PCR(Polymerase Chain Reaction)という方法を使って少量のサンプルから比較的簡単に反復数を検出することができるSTRがよく使われます。これらの変異情報は子孫に遺伝するので、適当な場所や組み合わせを選ぶと個人や親子鑑別を行うことができます。
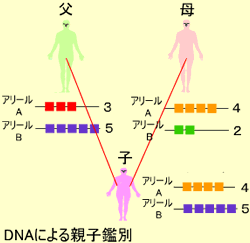
図のようにたとえばあるSTRで、子供が1対の各染色体上に4回と5回のリピートを持っていると父のアリールBからの5回のリピートと母のアリールAからの4回のリピートを受け継いだことがわかります。現在ではヒトだけでなく、他の動物(馬、豚、犬等)や植物にも同様の方法が応用されています。